

NÖRON DALLARINDAN AĞAÇLARA
Göle taş attığımız zaman etrafındaki suyun belli aralıklarla dalgalandığını görürüz. Bu dalgalanmaları inceleyerek suyun ve dolayısıyla moleküllerinin karakteristik özellikleri, kütleleri, yoğunlukları ve hızları hakkında bilgi sahibi olabiliriz. Chettih ve Harvey, fare beyinlerindeki birincil görsel korteksin mekanizmasını çözümlemek için benzer bir yaklaşım izliyorlar. Bu noktada göle attığımız taşı ilk nöron aktivitesine ve çevresinde kendisine göre daha geniş yayılan dalgaları da nöron ağlarına benzetebiliriz.
Yazarlar, her bir nöronun komşu nöronlar üzerindeki aktivitesini doğrudan ölçmek için “etki haritalaması” adını verdikleri bir yöntem kullandılar. Fareler, nöronlarındaki iyon kanallarını ışığa duyarlı hale getirebilmek için genetik olarak değiştirildi. Böylece ışığı nöral aktiviteyi kontrol etmek için kullandılar(optogenetik).Farelerde ayrıca nöronun elektriksel aktivitesini gösteren bir diğer gösterge olarak da kalsiyum konsantrasyonunu ölçmemizi sağlayacak floresan indikatörü eksprese edildi(calcium imaging tekniği). Optogenetik stimulasyon, hedef nöronlarda birçok aksiyon potansiyelini tetikledi. Dolayısıyla Chettih ve Harvey hedef nöronun çevresindeki nöronların etkilenmemesi için calcium imaging tekniğini kullandı. Çünkü hedefleri çevresel uyarıları ortadan kaldırarak tek bir nöronun diğerleri üzerine etkisini gözlemlemekti.
Bir nörondaki birkaç aksiyon potansiyeli iç içe geçmiş oldukça karmaşık nöron ağı için çöldeki bir kum tanesi gibidir. Ancak yazarlar, hedef ve komşu nöronlar arasındaki mesafeyi ve fonksiyonel özelliklerini(yani görsel uyarılara benzer veya farklı şekillerde cevap verip vermediklerini) göz önüne aldıklarında anlamlı bir tablo ortaya çıktı. Onların ortaya koyduğu “işlevsel bağlanabilirlik modelleri” bize birincil görsel kortekste gerçekleşen temel hesaplamalarla ilgili yol gösteriyor.
Son 15 yılda, hayvanlardaki tek nöron üzerindeki çalışmalar, bireysel nöronların ağlarına nasıl bağlandıklarını ve ağların genel işlevlerini anlamamıza katkı sağlamıştır. Bu çalışmaların çoğu sinaps adı verilen nöronlar arasındaki bağlantıların bol ama genel olarak zayıf olduğu korteksin sinir devrelerine odaklanmıştır. Ayrıca deneyler tek bir nöronun uyarılmasının davranışsal sonuçları olabileceğini göstermiştir.
Örneğin, motor korteksteki tek bir nöronun elektriksel olarak uyarılması farelerde bıyık hareketine neden olabilir. Sıçanlarda duyusal bilgilerin işlenmesinde yer alan barrel korteksindeki farklı nöronların benzer şekillerde uyarılması dokunma algısı ile ilişkili yalama davranışını tetikleyebilir. Tuhaf bir şekilde kortikal ağlar, uyarılmış nöronların hepsinin aktivasyonuna duyarlı olmadığı görülmüştür. Bu bulgu için Chettih ve Harvey bir açıklama getirmiştir.
Etki-haritalama yaklaşımı, elektrofizyoloji kullanarak çalışılanlardan daha büyük bir nöron popülasyonunun analiz edilmesini mümkün kılar. Daha da önemlisi yazarların nöronal etkileri, uyarılan nöronların tepkileri ile etkilediği komşu nöronların etkilerini aralarındaki benzerlik derecesine göre kategorize etmelerini sağlamıştır. Özellikle optogenetikle elde edilen nöronların incelikli bir şekilde uyarılması güçlü ve pek çok nöronu etkileyecek bir tepki oluşturması mümkün değildir. Bu yaklaşım, kortikal devrenin temel hesaplamasını diğer yaklaşımlardan daha iyi yansıtabilir.
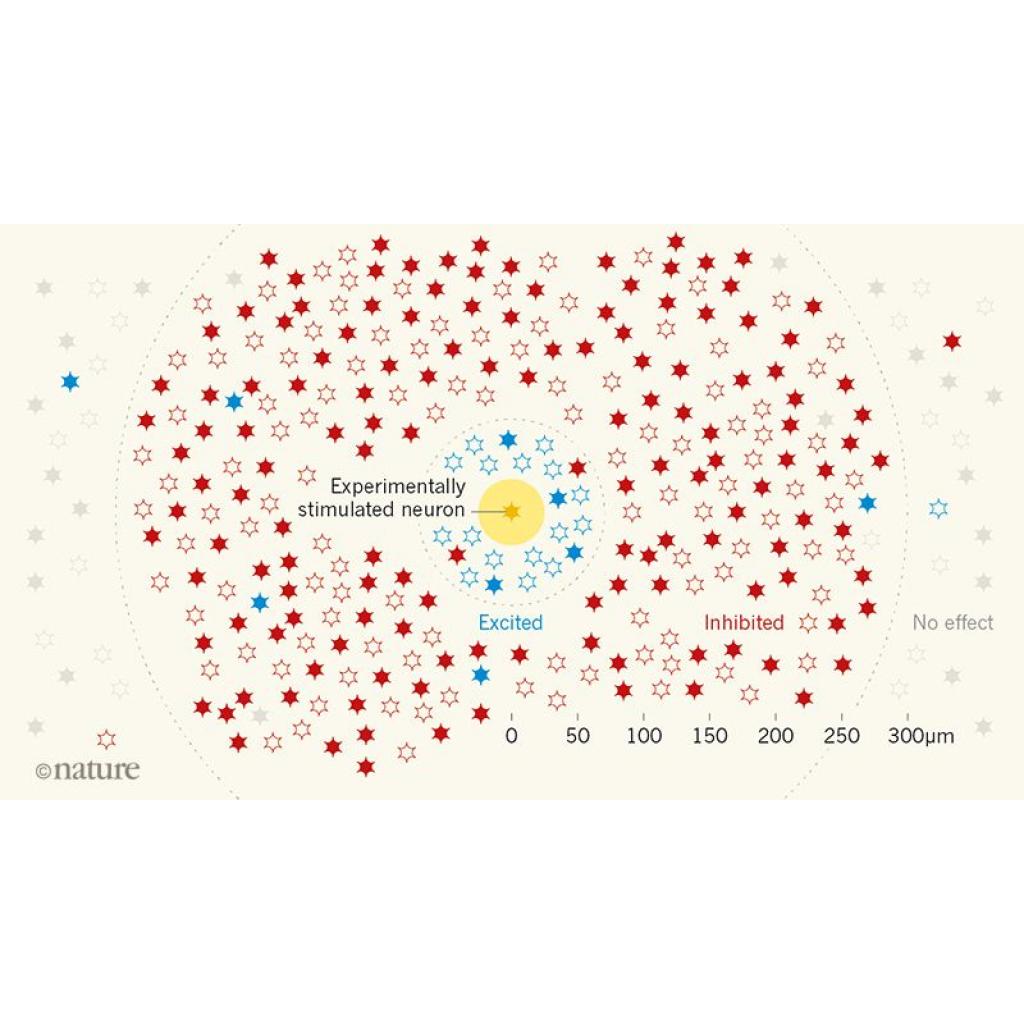
Şekil1: Tek bir nöronun çevresel ağa etkisi. Fare görme korteksindeki ağ modeli gösterilmiştir. Tek bir nöronun(sarı) deneysel olarak uyarılmasının ağırlıklı olarak komşu nöronların aktivitesinin inhibe edilmesine(kırmızı) neden olduğu bulunmuştur. Stimülasyon, kısa mesafelerde(25-70 µm) uyarıcı bir etkiye(mavi) sahip olsa da uzak mesafedeki(>300 µm) nöronlara(gri) neredeyse hiçbir etkisi olmamıştır. İçi boyanmış yıldızlar hedef nöronun görsel uyarana verdiği tepkiye benzer tepkiler verecek şekilde ayarlanmış nöronları temsil ederken içleri boyanmamış yıldızlar farklı şekilde ayarlanmış nöronları temsil eder. Nöral aktivetinin inhibisyona uğradığı alanda(70-300 µm), hedef nöronla benzerlik gösteren nöronlar, farklı olanlara göre daha güçlü bir şekilde inhibe edilmiştir. “Benzeri baskılama sistemi”(feature competition), yanıtları hedef nörona benzeyen az sayıda nörondan oluşan seyrek dağılmış aktifleştirme kümeleriyle(feature amplification) denge halindedir. Bu iki işlem uyaran seçiciliğini arttırır ve uyarının doğru işlenmesini sağlar.
Chettih ve Harvey ağırlıklı olarak hedef nöron etrafındaki nöral aktivitenin baskılandığını gözlemlemişlerdir(Şekil 1). Ayrıca bir nöronun çevresindeki nöronlarla etkileşiminin mesafeyle değiştiğini gördüler: Kısa mesafedeki küçük bir nöron popülasyonu üzerindeki etki(25-70 µm), orta mesafedeki inhibitör etki(70-300 µm), uzak mesafede çoğu komşu nöronu etkileyen diğerlerine göre daha az etki(>300 µm).
Yazarların daha sonraki detaylı araştırmaları, genel bir nöral aktivite inhibisyonundan çok daha karmaşık bir model ortaya koydu. Nöronların diğer nöronlar üzerindeki etkisinin derecesinin oryantasyon ve zamansal frekans gibi görsel uyaranların bazı özelliklerine nasıl cevap verdikleriyle ilgili olduğunu buldular. Bir nöron aktive edildiğinde, o nöronun benzer özelliklerine cevap verecek şekilde ayarlanmış nöronlar, farklı cevap vermek üzere ayarlanmış nöronlardan daha kuvvetli bir şekilde baskılanmıştır. Bu olgunun nöronların arasındaki uzaklığa bağlı olmaksızın geçerli olduğu görüldü. Bununla birlikte hedef nöronlarla benzer şekilde ayarlanmış seyrek dağılım gösteren nöron grubu ve bu grubun uyarıya güçlü bir şekilde tepki verdiği keşfedildi(Şekil1). Bu nöronların yanıt verme mekanizması da zamansal olarak hedef nöronunkiyle bağlantılıydı(anlık elektriksel aktiviteleri birbirlerine çok yakındı).
Yazarların bulguları, merkezi sinir sisteminin karmaşık ağının iki paralel hesaplama yönteminin dengelenmesiyle anlaşılabildiğini gösteriyor. Benzer ayarlara sahip nöronların arasındaki tek taraflı baskın inhibitör etkiyle(feature competition) iyi bağlanmış nöronların seyrek dağılım gösteren uyarıcı etkileri(feature amplification) arasında bir mücadele vardır. Model, inhibitör etkinin fazla uyarıları nasıl azalttığını ve anlamlı sinyallerin kalabalık uyaran grubundan nasıl ayıklandığını göstermekle birlikte; uyarıyı arttıran etkinin ise seçilen nöronların uyaran hakkında etkili bir şekilde bilgi taşıma kapasitesini arttırdığını açıklıyor.
Chettih ve Harvey’in vardığı sonuçlar pek çok soruyu da beraberinde getiriyor. Feature competition hücresel boyutta incelenebilir mi, işleyişi nasıldır? İnhibitör internöronlar diğer nöronların aktivitesini baskılamak için bölgesel olarak etki ederler ve farklı internöron tiplerinin uyarılara farklı cevapları vardır. Örneğin, bir nöronda beşten fazla aksiyon potansiyelinin görünmesi hedef nörona komşu olup somatostatin eksprese eden internöronların(SOM+) yaklaşık %30’unda ani bir tepki oluştururken; parvoalbumin eksprese eden internöronlarda(PV+) bir değişim gözlemlenmemiştir. Bunun tersini kanıtlayan bulgular da mevcuttur. Chettihve Harvey yaklaşık olarak hedef nöronda optogenetiği kullanarak altı aksiyon potansiyeli oluşturdu. Böyle bir uyarı SOM+ internöronlarını PV+ internöronlarına göre daha yüksek bir ihtimalle aktifleştirebilir. Özellikle SOM+ internöronları diğerlerine(PV+) göre daha seçici bir şekilde ayarlanmıştır. Ayarlanmış inhibitör nöronların özellikle güçlendirilmesi feature competition etkisini gözlemlerken bizi yanlış bir sonuca sürükleyebilir.
Beyin uyaranı algılarken hayvanın davranışsal durumundan nasıl etkilenir? Kortikal ağ farklı davranışsal ve çevresel faktörler için nöronal tepkileri etkileyecek farklı algılama sistemleri mi geliştiriyor? Sinir ağındaki dallanmalar daha büyük bir uyaran karşısında çarpıcı bir şekilde değişebilir mi?
Yararlanılan Kaynaklar
1. Chettih, S. N. & Harvey, C. D. Nature 567, 334–340 (2019).
2. London, M., Roth, A., Beeren, L., Häusser, M. & Latham, P. E. Nature 466, 123–127 (2010).
3. Kwan, A. C. & Dan, Y. Curr. Biol. 22, 1459–1467 (2012).
4. Jouhanneau, J.-S., Kremkow, J. & Poulet, J. F. A. Nature Commun.9, 1540 (2018).
5. Brecht, M. Curr. Biol. 22, R633–R635 (2012).
6. Brecht, M., Schneider, M., Sakmann, B. & Margrie, T. W. Nature427, 704–710 (2004).
7. Houwelling, A. R. & Brecht, M. Nature 451, 65–68 (2008).
8. Ko, H. et al. Nature 496, 96–100 (2013).
9. Li, C. T., Poo, M. & Dan, Y. Science 324, 643–646 (2009).
10. Ma, W. et al. J. Neurosci. 30, 14371–14379 (2010).